|
Dipartimento di Scienze Animali, Vegetali e dell'Ambiente, Università degli Studi del Molise, Via F. De Sanctis, 86100 Campobasso. E-mail: lima@unimol.it
RIASSUNTO
Più del 90% delle specie vegetali terrestri, ivi incluse le piante di interesse agrario presentano micorrize. Negli ultimi anni grazie all’accresciuto interesse su questi rapporti simbiotici e alla disponibilità di nuove e raffinate tecniche di indagine genetico-molecolare e biochimiche le conoscenze su queste associazioni pianta-fungo hanno subito una considerevole espansione. Gli effetti benefici sullo stato nutrizionale, sulla tolleranza agli stress abiotici e biotici delle piante nonché le ripercussioni positive sull’ambiente hanno stimolato l’attenzione sulle possibilità applicative delle micorrize nel settore agrario e forestale.
ABSTRACT
More than 90% of terrestrial plants, including agricultural and forestry species, are associated with root-colonizing fungi, establishing a permanent and intimate mutualistic symbiosis, called mycorrhiza. During the last years, due to both increasing interest on mycorrhizae and availability of reliable and advanced genetic-molecular and biochemical tools, the knowledge about the role of mycorrhizae in the environment has significantly expanded. The beneficial effects of mycorrhizae on nutritional condition and tolerance of plants to abiotic and biotic stresses have stimulated the interest of researchers and plant producers on possible applications of these symbiotic fungi for agricultural and forest purposes.
Parole chiave: funghi simbionti, micologia, stress ambientali, fitopatogeni, bioprotezione.
Key words: symbiotic fungi, mycology, environmental stress, plant pathogens, bio-protection.
Le simbiosi micorriziche
L’associazione tra le radici di una pianta superiore e un fungo, definita micorriza, è la forma di simbiosi mutualistica più diffusa in natura e forse anche la più remota. Infatti, più del 90% delle specie vegetali degli ambienti naturali sono micorrizate e resti fossili evidenziano come le micorrize erano già presenti diversi milioni di anni fa quando i primi vegetali fecero la loro comparsa sulle terre emerse (Simon et al., 1993; Koide e Mosse, 2004). Tuttavia, la scoperta e la comprensione del fenomeno della micorrizazione da parte dell’uomo risale a poco oltre 100 fa. Il termine “mycorrhiza” fu, infatti, coniato per la prima volta da Bernhardt Frank (1885) il quale segnalò speciali strutture nelle radici delle piante. Lo stesso Frank oltre a descrivere la morfologia intuì anche il ruolo fisiologico delle stesse micorrizze (Frank, 1888).
I funghi sono organismi eterotrofi, privi di clorofilla e pertanto per le loro funzioni vitali e riproduttive devono attingere sostanza organica elaborata da altri organismi. Nelle micorrize, il fungo preleva dalla pianta buona parte, se non tutti, i composti organici e rende maggiormente disponibili per la pianta l'acqua ed i sali minerali e altre sostanze utili che riesce ad assorbire dal terreno. Il rapporto simbiontico si realizza a livello degli apici radicali. Le radici interessate dalla simbiosi sono generalmente quelle secondarie, ricche di capillizio e specializzate nell'assorbimento delle sostanze nutritive dal terreno.
In base al tipo di interazione tra il fungo e la radice è possibile distinguere differenti tipologie di micorrize (Fig. 1), ma in funzione del tipo di interazione morfo-fisiologica tra radice e fungo esse possono essere collocate in due gruppi: le ectomicorrize e le endomicorrize (Deacon, 2000).
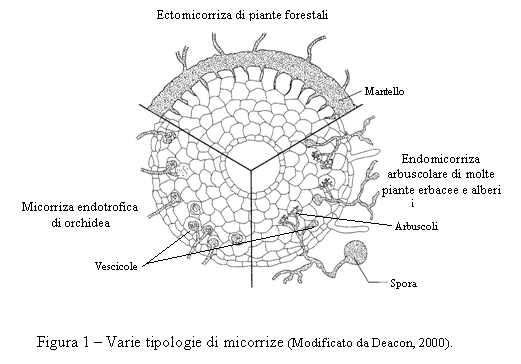
Nelle ectomicorrize le ife del fungo rimangono esterne all'apice radicale della pianta, avvolgendolo completamente fino a formare un mantello, una sorta di guaina di alcuni strati di cellule, che prende il nome di micoclena; lo scambio di materiale nutritivo tra fungo e pianta avviene a livello di alcune ife che si insinuano tra le cellule della corteccia radicale e formano una massa micelica reticolata che prende il nome di reticolo di Hartig. A seconda dell’ospite il reticolo può essere più o meno sviluppato, e raggiungere il cilindro centrale (conifere) oppure limitarsi ai primi strati cellulari della corteccia (latifoglie). Dal mantello si diparte una fitta rete di ife esterne e cordoni miceliari che si estendono notevolmente nel suolo circostante e operano, insieme ad altre ife, l'assorbimento dell'acqua e dei sali minerali presenti nel terreno e, attraverso le ife del reticolo di Hartig, li trasferiscono alla pianta ospite. Le ife esterne, in condizioni appropriate possono produrre le strutture riproduttive (carpofori). Le ectomicorrize sono tipicamente diffuse fra le piante forestali e vedono coinvolte numerose specie di funghi ascomiceti e basidiomiceti caratterizzati dalla presenza di carpofori. Tra questi vi sono i tartufi (Tuber spp.), i porcini (Boletus spp.) ed altre note specie di funghi macroscopici. Nelle simbiosi ectomicorizziche, la formazione dei corpi fruttiferi nel terreno si realizza quando si verificano le seguenti condizioni: - la pianta simbionte ha raggiunto la maturità fisiologica; - nel terreno c'è una sufficiente carica di micorrize; - le condizioni ecologiche sono favorevoli. Solo allora le ife bloccano il loro accrescimento ed iniziano a formare il carpoforo. Le primissime fasi di formazione del carpoforo sono poco note, ma si ipotizza che inizialmente questo sia costituito da un intreccio globoso di ife (primordio o abbozzo del carpoforo), sviluppatesi a partire dalle micorrize.
Le endomicorrize sono quelle a più ampia diffusione (anche tra le specie erbacee). In queste micorrize, le spore fungine, stimolate dalla presenza di essudati radicali dell’ospite, germinano nel terreno e le ife fungine si sviluppano fino a raggiungere la radice. Il fungo si diffonde nei tessuti corticali sia intercellularmente sia direttamente nelle cellule, ma senza mai raggiungere il cilindro centrale e le cellule dell’apice radicale e senza formare alcuna struttura esterna. La presenza di questo tipo di micorrize, infatti, non è rilevabile ad occhio nudo, ma soltanto al microscopio, sezionando la radice. Il fungo si evidenzia in quanto all'interno delle cellule radicali le sue ife si diramano formando strutture ramificate molto sottili, gli arbuscoli, responsabili degli scambi nutrizionali tra i due simbionti. Gli arbuscoli hanno vita breve: dopo pochi giorni, infatti, degenerano. Altre strutture talvolta prodotte dalle ife fungine di talune micorrize sono le vescicole, rigonfiamenti tondeggianti inter o intracellulari, ricche di lipidi, con probabile funzione di riserva (Fig. 1).
Le endomicorrize arbuscolari (AM) rappresentano il più comune tipo di micorrizia e si riscontrano in molte specie vegetali, sia coltivate sia spontanee (erbacee, ortive, da frutto, forestali, tropicali, orchidee e perfino pteridofite e briofite). Le AM sono prodotte da numerose specie di funghi biotrofi appartenenti all’ordine delle Glomales (Zygomiceti). A parte alcune eccezioni, generalmente non vi è specificità tra fungo e pianta ospite. Quindi, ad esempio, un fungo simbionte capace di svilupparsi su una pianta forestale potrà essere in grado di farlo anche su una pianta di interesse agrario e viceversa.
Micorrize, pianta e ambiente
Nonostante i numerosi studi degli ultimi anni, i meccanismi dell’interazione fisiologica tra funghi simbionti e radici vegetali sono ancora poco noti (Hodge, 2000; Johansson et al., 2004). E’ indubbio, comunque, che la simbiosi micorrizica comporta notevoli vantaggi sia per la pianta microrrizata sia per il fungo. Negli ambienti fortemente antropizzati (ad es. campi coltivati e verde urbano) le micorrize sono spesso assenti oppure presenti in forma molto ridotta, molto probabilmente a causa dei vari fattori di inquinamento chimico del suolo. La pianta micorizzata presenta un migliore assorbimento dell'acqua ed una più efficiente assunzione degli elementi minerali. Infatti, lo sviluppo considerevole delle ife fungine permette alle radici di esplorare ed acquisire acqua e sali minerali da un volume di suolo notevolmente maggiore, raggiungendo zone di assorbimento normalmente al di fuori dalla portata della radice stessa. Le micorrize, inoltre, sono in grado di solubilizzare, assorbire e quindi rendere disponibili per la pianta, forme organiche o minerali solitamente presenti nel suolo come composti insolubili, cambiando radicalmente il conto agronomico della disponibilità degli elementi nutritivi nel terreno. La micorrizazione, inoltre, comporta un incremento nel ritorno di sostanza organica al suolo accompagnato anche da un maggior rilascio di azoto, fosforo e potassio. La migliore nutrizione minerale (soprattutto fosfatica) si traduce in una maggiore crescita della pianta, maggiormente evidente nei terreni con scarso contenuto di elementi minerali. Le piante micorrizate sono spesso più competitive e meglio tollerano le condizioni di stress rispetto alle piante non micorrizate. La pianta, per contro, cede al fungo zuccheri semplici prodotti attraverso la fotosintesi e composti organici elaborati dalla cellula vegetale.
Il fungo, grazie alla simbiosi, è in grado di completare il proprio ciclo vitale, e nel caso delle ectomicorrize, di formare i corpi fruttiferi. Dalle micorrize si sviluppano poi nuove ife che vanno a colonizzare il terreno circostante ed anche le nuove radichette emesse dalla pianta o quelle di altre piante vicine.
Il ciclo di attività delle micorrize segue quello della pianta: in primavera, con la ripresa vegetativa, le micorrize riprendono a crescere e continuano per tutta l'estate se le condizioni del terreno si mantengono favorevoli. All'arrivo dell'inverno le micorrize riducono l'attività metabolica per affrontare la stagione avversa e, quelle che riescono a superarla, alla primavera successiva riprenderanno il loro sviluppo.
Il potenziale d’inoculo può essere ridotto da certe pratiche agricole, come la fertilizzazione e le lavorazioni profonde, oppure, nei terreni incolti, dall’assenza di piante simbionti.
Tutti i funghi micorrizici svolgono un ruolo ecologico importantissimo nel riciclo degli elementi nutritivi presenti nel terreno perché, grazie all'assorbimento da essi mediato, riducono il rischio di perdite di percolazione. Nei suoli forestali, tipicamente poveri, questo fenomeno è di fondamentale importanza. La maggiore capacità di assorbimento delle piante micorrizate si traduce, praticamente, in uno sviluppo superiore rispetto a quelle non micorrizate, e rende conveniente l'impiego di piante preventivamente micorrizate nei rimboschimenti.
Nelle ectomicorrize la radice micorrizata appare profondamente trasformata, la crescita è bloccata e l’apice radicale si ramifica e le radici assumono un aspetto coralloide (Fig. 2). Il mantello fungino spinto dalle radichette secondarie non si rompe, ma si estende entrando in attiva proliferazione cellulare, inglobando le nuove radici laterali.
Nelle endomicorrize, sebbene la radice non subisca variazioni morfologiche così notevoli, l’apparato radicale risente della presenza del fungo, variando il grado di ramificazione e le dimensioni delle radici stesse, fino ad aumentare di centinaia di volte.

Figura 2 – Effetti morfologici della micorrizazione ectotrofica su radici di nocciolo. In A glomeruli di radici micorrizzate dal tipico aspetto coralloide; In B, particolare dell’ingrossamento e della singolare ramificazione tricotomica di un apice radicale.
Aspetti fitopatologici
Negli ultimi decenni, differenti studi sono stati orientati ad approfondire le conoscenze sulle interazioni tra funghi simbionti, piante, e fitopatogeni (Dehne, 1982; Hussey e Roncadorl, 1982; Jalali e Jalali, 1991; Sharma et al., 1992; Johansson et al., 2004). E’ ormai certo, che le micorrize, in particolare quelle arbuscolari (Arbuscular Mycorrhizae: AM), incrementano il benessere della pianta non solo attraverso il miglioramento dell’assunzione di nutrienti e della tolleranza allo stress, ma spesso anche attraverso induzione di resistenza alle malattie. Tuttavia, esistono anche casi ben documentati, riguardanti soprattutto patogeni fogliari, di aumentata suscettibilità alle malattie in piante micorizzate (Gernns et al., 2001) giustificabile, con ogni probabilità, dal maggior rigoglio vegetativo della pianta micorrizzata.
Integrandosi nel sistema radicale della pianta, le micorrize interagiscono anche con i microorganismi presenti nel suolo richiamati verso la radice dall’incremento di essudati radicali. Tali microrganismi, unitamente alle micorrize, possono profondamente influenzare sia la fisiologia e della pianta sia la colonizzazione da parte di patogeni ad habitat terricolo (soil-bone pathogens). Le micorrize arbuscolari sono in grado di modificare la composizione degli essudati, incrementare l’attività chitinolitica, l’efficienza fotosintetica e respiratoria.
Tra i meccanismi coinvolti nell’induzione di resistenza alle malattie da parte di funghi simbionti si segnalano a) l’esclusione del patogeno, a causa ad esempio delle barriere fisiche costituite dalla presenza delle strutture micorriziali, b) la lignificazione delle pareti delle cellule radicali, c) le variazioni indotte nell’essudazione radicale, d) la formazione de novo di composti a basso peso molecolare dotati di attività inibitoria nei confronti dei fitopatogeni.
Per queste loro caratteristiche i funghi AM presentano notevoli potenzialità ad essere utilizzati come agenti di lotta biologica in Patologia vegetale (Johansson et al., 2004). Al riguardo, specifiche ricerche hanno evidenziato come la micorrizazione artificiale di specie vegetali appare un’efficace strategia per il controllo di soil-borne pathogens ivi compresi organismi che risultano di difficile controllo perfino con mezzi chimici e fisici di lotta (Azcon-Aguilar e Barea, 1997; Cordier et al., 1997). Prodotti a base di specie fungine simbiotiche selezionate, moltiplicate e opportunamente formulate, sono già disponibili in commercio per la microrrizazione di piante di interesse agrario (piante ortive e da frutto), forestale e ornamentale. Nella figura 3, si riporta a titolo di esempio la confezione di uno di questi prodotti disponibili in commercio in Italia.
In conclusione, i funghi responsabili di simbiosi micorriziche presentano notevole interesse applicativo per i loro possibili molteplici impieghi. Nonostante i progressi della ricerca conseguiti in questi ultimi anni, ulteriori studi sembrano necessari per una migliore comprensione dei meccanismi coinvolti nelle complesse interazioni tra radici e funghi simbionti e tali conoscenze potranno senz’altro contribuire ad un notevole sviluppo nella utilizzazione e nell’ottimizzazione dell’impiego della micorriza per la tutela dell’ambiente.

BIBLIOGRAFIA
Azcon-Aguilar C. & J.M. Barea - 1997: Applying mycorrhiza biotechnology to horticulture: significance and potentials. Scientia Horticulturae 68: 1–24.
Cordier C., A. Trouvelot, S. Gianinazzi & V. Gianinazzi-Pearson – 1997: Arbuscular mycorrhiza technology applied to micropropagated Prunus avium and to protection against Phytophthora cinnamomi. Agronomie 17: 256-265.
Deacon J.W. - 2000: Micologia moderna. Calderini Edagricole, Bologna, 414 pp.
Dehne H.W. – 1982: Interaction between vesicular-arbuscular mycorrhizal fungi and plant pathogens. Phytopathology 72(8): 1115-1119.
Frank B. – 1885: Ueber die auf Wurzelsymbiose beruhende Ern¨ahrung gewisser B¨aume durch unterirdische Pilze. Ber. Dtsch. Bot. Ges. 3: 128-145.
Frank B. – 1888: Uber die physiologische Bedeutung der Mycorrhiza. Ber. Dtsch. Bot. Ges. 6: 248-269.
Gernns H., · H. von Alten & H.M. Poehling - 2001: Arbuscular mycorrhiza increased the activity of a biotrophic leaf pathogen – is a compensation possible? Mycorrhiza 11: 237-243.
Hodge A. - 2000: Microbial ecology of the arbuscular mycorrhiza. FEMS Microbiology Ecology 32: 91-96.
Hussey R.S. & R.W. Roncadorl – 1982: Vesicular-arbuscular mycorrhizae may limit nematode activity and improve plant growth. Plant Disease 66(1): 9-14.
Jalali B.L. & I. Jalali – 1991: Mycorrhiza in plant disease control. In: Arora K., B. Rai, K.G. Mukerji & G.R. Knudsen (eds), Handbook of applied mycology. Dekker, New York, pp 131–154.
Johansson J.F., L.R. Paul & R.D. Finlay - 2004: Microbial interactions in the mycorrhizosphere and their significance for sustainable agriculture. FEMS Microbiology Ecology 48: 1-13.
Koide R.T. & B. Mosse - 2004: A history of research on arbuscular mycorrhiza. Mycorrhiza 14: 145-163.
Sharma A.K., B.N. Johri & S. Gianinazzi - 1992: Vesicular-arbuscular mycorrhizae in relation to plant disease 8(6): 559-563.
Simon L., J. Bousquet, R.C. Levesque & M. Lalonde - 1993: Origin and diversification of endomycorrhizal fungi and coincidence with vascular land plants. Nature 363: 67-69.
|